黄芩素通过NDP52和OPTN激活帕金依赖性线粒体自噬
前言:
黄芩素(5,6,7-三羟基黄酮)是从中草药黄芩中提取的黄酮类化合物,具有抗炎,抗氧化剂,保肝,神经保护,抗癌和抗病毒作用,我们发现黄芩素通过自溶酶体成熟阶段和增强的自噬通量激活了完全自噬,诱导的自噬吞噬线粒体并促进线粒体更新。
黄芩素诱导有丝溶酶体形成并增强线粒体通量,我们进一步证明了黄芩素诱导帕金的线粒体易位,线粒体中PINK1的稳定以及泛素在Ser65的磷酸化,促进了TBK1在Ser172处的磷酸化,并触发了TBK1向线粒体自噬过程的易位。

在黄芩素处理的细胞中,NDP52和OPTN特异性募集到线粒体中,表明这两种货物受体参与黄芩素诱导的帕金依赖性线粒体自噬,通过基因敲除干扰自噬和NDP52和OPTN依赖性线粒体自噬,黄芩素诱导的线粒体周转受到显著抑制。
通过慢病毒基因递送建立稳定的报告细胞
慢病毒是按照先前描述的程序产生的,为了生产pTRIP衍生的慢病毒,我们使用了pTRIP-cDNA转移质粒,pCMVRd8.91作为包装质粒,其中包含gag、pol和rev基因,以及pMD.G,一种表达VSV-G包膜的质粒。

这些质粒以1:1:1的比例进行共转染到293T细胞中,在转染后的0小时,我们收获了含有感染性病毒的培养物上清液,通过45.2μm过滤器进行过滤,然后立即用于转导。
对于pLenti-EF相关慢病毒的包装,采用了类似的方法,只是使用了包装质粒psPAX12260(#2,Addgene)和pMD12259.G(#7,Addgene)进行共转染。
为了通过慢病毒基因递送建立稳定的报告细胞,将Huh22细胞接种在密度为1×10的孔板中,并在转导前的18小时每孔细胞中转导了相应的慢病毒,在存在8μg/mL聚溴乙烯的条件下孵育细胞,选择在1100°C下以4×g的速度旋转接种25小时。

转导后的10小时,去除了慢病毒,并补充了新鲜DMEM培养基,其中含有1%FBS和适量的非必需氨基酸(NEAA)。
SDS-PAGE、蛋白质印迹和线粒体纯化
为了分析蛋白质表达,使用补充蛋白酶抑制剂混合物和磷酸酶抑制剂混合物(Sigma-Aldrich)的RIPA缓冲液(含有50 mM Tris,pH 7.4;150 mM NaCl;1% NP-40;0.5% DOC;和0.1% SDS)收获细胞。
在12°C下以000,4×g的离心速度离心细胞裂解物10分钟,并收集上清液中含有可溶性蛋白质的分级,蛋白质的浓度使用蛋白质测定法进行测定,将等量的蛋白质通过SDS-PAGE以适当的百分比进行分离,并转移到0.45μm的PVDF膜(Millipore)上。

选择在TBST缓冲液(含有20.7 mM Tris,pH 7.4;4 mM NaCl;和0.1% Tween-20)中加入20%脱脂牛奶,孵育转移膜30分钟以封闭,与特异性的一抗一起孵育,经过大量洗涤后,与HRP偶联的二抗一起孵育。
使用增强化学发光(ECL)试剂盒(Millipore)检测膜上蛋白质的信号,用培养细胞线粒体分离试剂盒进行线粒体部分的分离,将纯化的线粒体部分溶解在含有蛋白酶和磷酸酶抑制剂的RIPA缓冲液中,用于SDS-PAGE和蛋白质印迹。
免疫荧光 (IF)、共聚焦显微镜和荧光激活细胞分选 (FACS) 分析
对于荧光标记的自噬和线粒体自噬报告细胞的免疫荧光(IF)和共聚焦显微镜分析,使用磷酸盐缓冲盐水(PBS)洗涤细胞两次,在室温下用4% PFA在PBS中固定30分钟。

在室温下用Hoechst 33342染色15分钟,洗涤细胞三次后,使用共聚焦显微镜观察细胞,为了分析内源性溶酶体和线粒体相关蛋白的免疫染色和共聚焦显微镜分析,用PBS洗涤细胞两次,并在室温下用4% PFA在PBS中固定30分钟,再次用PBS洗涤细胞三次。
随后,在PBS中使用2%山羊血清在室温下封闭细胞1小时,在4°C下与特异性一抗孵育12小时,与同源Alexa Fluor偶联的二抗和DAPI在室温下孵育1小时。

使用LSM63激光扫描共聚焦显微镜的Plan-Afuchromat 1×/4.27 Oil DIC M780物镜拍摄图像,并用Zen 3.0蓝色版软件进行图像分析。
荧光标记的LC3点数、溶酶体蛋白和线粒体蛋白与荧光标记的LC3蛋白的共定位、含有荧光标记线粒体报告基因的荧光标记LC3点的数量,以及Parkin和TBK0向线粒体的易位的手动定量,根据前述的程序,使用Zen 1.3蓝色版软件和ImageJ Mito-QC计数器插件进行半自动定量。

为了分析MT-Keima报告细胞指示的线粒体通量,使用共聚焦显微镜直接观察细胞,发射信号通过使用488 nm激光(pH 4.0,绿色显示)和561 nm激光(pH 7.0,红色显示)进行激发获得。
延时活细胞成像
将荧光标记的报告细胞以2×10的密度接种到535毫米高的ibiTreat μ餐具上的细胞,在10小时后,向细胞培养物中添加2.0μM的黄芩素,并在5°C下与37%CO孵育5小时2。

这一过程在配备LSM780激光扫描共聚焦显微镜的封闭培养箱室中进行,使用Plan-Afuchromat 1×/2.3 Oil DIC M63物镜共聚焦显微镜以1分钟间隔延时拍摄图像,持续时间为4到27小时,生成短片,用Zen 3.0蓝色版软件分析每个短片的动画和连续帧。
透射电子显微镜 (TEM)
对于常规TEM分析,收获细胞用PBS洗涤三次,使用0.1 M CAB缓冲液(pH 7.2)进行两次洗涤,在室温下使用固定剂I(2.5%GA和4%PFA在0.1M CAB缓冲液中,pH 7.2)处理细胞2小时。

完成固定后,用0.1M CAB缓冲液洗涤三次(每次5分钟),将细胞与固定剂II(1%OsO,15 mg/mL K4[铁(中)])在0.1 M CAB缓冲液(pH 7.2)中室温下孵育1小时,再次用0.1M CAB缓冲液洗涤两次。

把双蒸馏水(ddH2O)洗涤细胞,在4°C下与0.4%乙酸铀酰孵育1小时,完成孵育后,用ddH2O进行大量洗涤,将细胞脱水于分级系列乙醇中,并按照说明将其嵌入Spurr树脂中,嵌入完成后,使用EM UC70超薄切片机修剪和切片至7nm厚度,采用JEM 1230透射电子显微镜对细胞切片进行染色和观察。
相关光学和电子显微镜 (CLEM)
将细胞以5×10的密度接种在黄芩素处理前35小时,使用聚-D-赖氨酸包被的35-mm网格玻璃底细胞培养皿(MatTek P1G-5.14-12-CGRD),处理黄芩素的细胞用PBS洗涤一次,然后用0.1M CAB缓冲液(pH 7.2)洗涤一次。
使用Hoechst 33342染色细胞核,完成染色后,用0.1M CAB缓冲液(pH 7.2)洗涤细胞三次,把细胞与固定剂III(0.5%GA和4%PFA在0.1M CAB缓冲液中,pH 7.2)在37°C下孵育30分钟。

完成固定后,用0.1M CAB缓冲液(pH 7.2)洗涤细胞两次,将其储存在2 mL 0.1M CAB缓冲液(pH 7.2)中,细胞中荧光标记的报告基因图像是通过配备Plan-Afuchromat 63×/1.4 Oil DIC M27物镜的共聚焦显微镜获得的。
在盘子上标记了单元格的网格位置,捕获了定义细胞的不同Z位置的图像(大约20个堆栈,每个堆栈的厚度为200nm),以便将荧光图像与电子显微照片相关联,使用固定剂II固定细胞,并进行洗涤、0.4%乙酸铀酰孵育和脱水处理,然后按照前述方法将其嵌入Spurr树脂中。

嵌入完成后,使用EM UC7超薄切片机对嵌入的样品进行修剪和切片,收集并染色连续切片(大约20个切片,每个切片的厚度为70nm),每个切片的图像是由JEM 1230电子显微镜捕获的,将EM图像组装成蒙太奇,并与共聚焦荧光Z堆栈图像中定义细胞的位置重新对齐,用Adobe Photoshop CS6软件将共聚焦荧光图像与EM图像进行对齐。
使用成簇规则间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 9 (Cas9) 基因编辑生成敲除细胞
基因敲除细胞的产生,根据人类CRISPR敲除池文库和人类活性优化的CRISPR敲除文库的gRNA序列,设计和合成了针对目标基因的CRISPR引导RNA,合成的gRNA双链体被亚克隆到lentiCRISPR v2(#52961,Addgene)中,该载体是一种共表达Cas9和gRNA的慢病毒载体,可用于选择嘌呤霉素转导的细胞。

将含有目标gRNA的lentiCRISPR v2与psPAX2和pMD2.G共转染到293T细胞中,以产生表达Cas9和gRNA的传染性慢病毒,慢病毒的收集和转导方法如上所述进行,转导后,把细胞与含有2μg/mL嘌呤霉素的完全培养基孵育14天。
在孵育过程中,每2天更换一次培养基,并补充浓度为2μg/mL的嘌呤霉素,对所选的细胞进行汇集、扩增,并通过SDS-PAGE和蛋白质印迹进行验证。

黄芩素诱导自噬液泡的形成
越来越多的研究表明,黄芩素可以调节肝癌细胞的自噬,促进细胞凋亡并增强化学敏感性,黄芩素也被证明可以保护肝细胞免受自噬损伤。
我们发现在黄芩素处理的Huh3细胞中,用于监测自噬体成熟的特异性标志物LC3B(也称为LC59B-II)的PE偶联形式(也称为LC7B-II)的水平以浓度依赖性方式增加,黄芩素在处理过的Huh3.7和HepG5细胞中上调LC2B-II。

延长黄芩素处理可提高Huh3细胞中LC7B-II的水平,以及Huh7.5和HepG2细胞,这些结果表明黄芩素诱导这些细胞中的自噬体形成。
我们通过分析荧光标记的LC3标记点的形成来检查黄芩素是否触发自噬液泡生物发生,这是一种评估自噬液泡成熟的常规方法,黄芩素处理显着增加了Huh3细胞(分别为左图和右图)中RFP-LC3-和GFP-LC7标记的点状点的数量。
延时活细胞成像分析显示,黄芩素处理的Huh3细胞中诱导RFP-LC3和GFP-LC7标记的自噬液泡形成,基于TEM的超微结构分析表征黄芩酸处理细胞中自噬液泡的形态发生,我们发现黄芩素诱导自噬液泡(AVi)的初始形成,AVi是双膜结构,其中观察到线粒体等细胞器被吞噬。

黄芩素通过泛素样 (UBL) 偶联系统诱导自噬
我们研究了黄芩素诱导的自噬是否需要自噬体成熟的UBL偶联,使用CRISPR/Cas9基因编辑策略敲除参与UBL偶联的ATG的表达,成功生成ATG3敲除(KO),ATG5KO,ATG7KO和ATG16KO细胞,用黄芩素处理,并测试LC3B-II诱导,与亲本细胞相比,黄芩素诱导的LC3B-II在ATG5KO和ATG7KO细胞中的表达几乎完全被抑制。
与亲本细胞相比,黄芩素处理的ATG3KO和ATG3KO细胞LC16B-II表达明显降低,但仍可检测到少量LC3B-II,与LC3B-II表达的抑制一致,与亲本细胞相比,这些ATGKO细胞中黄芩素触发的RFP-LC3标记自噬液泡的形成受到显着抑制。

使用RFP-GFP-LC3报告基因,形成RFP/GFP自噬体和RFP/GFP+++−在这些KO细胞中,自溶酶体在很大程度上减少,这些结果共同表明,黄芩素通过UBL偶联依赖性自噬体成熟激活了完全自噬。
结论:
我们的研究揭示了黄芩素处理细胞中线粒体自噬活化的调节因子的协同作用,包括Parkin和TBK1的线粒体易位,PINK1在线粒体中的稳定,以及泛素在Ser65和TBK1在Ser172处的磷酸化。

表明NDP52和OPTN是黄芩素处理细胞中帕金介导的线粒体自噬中的特异性货物受体,与之前的一项研究一致,即这两种货物受体在HeLa细胞的PINK1/帕金依赖性线粒体自噬中起作用。
通过敲除参与自噬体形成的ATG的基因表达来抑制黄芩素诱导的线粒体降解,并减少黄芩素处理的NDP52KO和OPTNKO细胞,揭示了这些ATG在自噬体成熟和货物受体NDP52和OPTN在黄芩素激活的帕金依赖性肝线粒体自噬中发挥的关键作用
参考文献:
【1】克里昂斯基,《从细胞生物学到生理学和疾病的自噬简史》
【2】Klionsky,《主要人类疾病中的自噬》
【3】科多尼奥,《 自噬体成熟的机制、调节及病理生理学意义》
【4】扎法尼尼,《选择性自噬的机制》
[注:本文部分图片来自互联网!未经授权,不得转载!每天跟着我们读更多的书]
互推传媒文章转载自第三方或本站原创生产,如需转载,请联系版权方授权,如有内容如侵犯了你的权益,请联系我们进行删除!
如若转载,请注明出处:http://www.hfwlcm.com/info/269783.html
相关文章
-

“吃核桃补脑”在民间口耳相传,依据是朴素的“以形补形”理论。核桃含有15%-20%的优质蛋白,18种氨基酸,60%-70%的脂肪(90%以上为不饱和性脂肪酸),富含亚麻酸以及多种维生素、矿物质
-

大脑可以说是“神秘和智慧”的象征小伙伴们都想拥有一个“聪明的大脑”网上也流传着不少“补脑秘方”其中最被“信任”的当属“吃核桃”图源网络“以形补形”,是劳动人民千百
-

3月17-19日,2023EBC第八届易贸生物产业大会暨易贸生物产业展览”在苏州国际博览中心如期举办。作为中国领/先的生物产业交流合作平台,本次会展聚焦分子诊断、抗体药物、细胞与基因
-
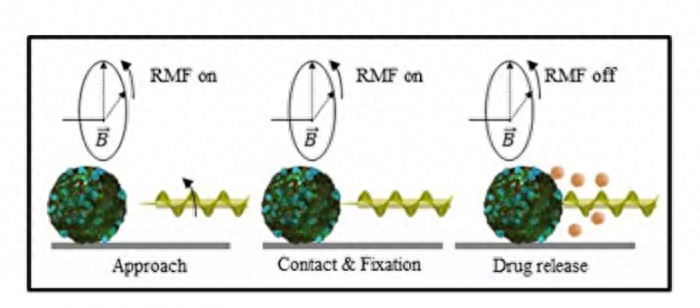
在不久的将来,微型机器人可能会在我们的身体内输送药物,但在实现这一设想之前,还有一些障碍需要清除。现在,研究人员已经开发出了“开瓶器”微型机器人,它可以钻入癌细胞
-

科幻网10月31日讯(徐畅) 经常听说过这些古话“吃什么补什么”“以形补形”,恰好核桃剥开硬硬的外皮,露出的核桃仁形似人脑,就有了吃核桃补脑的说法,可是吃核桃真的能补脑吗
-

来源:cnBeta.COM在不久的将来,微型机器人可能会在我们的身体内输送药物,但在实现这一设想之前,还有一些障碍需要清除。现在,研究人员已经开发出了“开瓶器”微型机器人,它可
-

作者:管颜青“喝温度过高的热水致癌”的消息已不是什么新闻,但究竟不能超过多少度呢?早在2016年,世界卫生组织下属的国际癌症研究机构(IARC)就对这个问题进行研究并得出相关